复能易锦提供的全部人类和小鼠ORF克隆及相关慢病毒和AAV病毒,所提供的载体质粒均采用毛细管电泳sanger全长测序验证,并承诺编码的氨基酸序列与NCBI数据库匹配。 All human and mice ORF clones and related Lentivirus, AAV particles and recombinant proteins provided by iGeneBio are fully sequenced using Sanger sequencing by capillary electrophoresis and Amino Acid Sequences are guaranteed to be matched with NCBI database.
Hasan, K.M., et al. (2023). Fatty Acid Excess Dysregulates CARF to Initiate the Development of Hepatic Steatosis. Cells doi: 10.3390/cells12071069 [AAVPrime™ AAV particles expressing CARF and AAV-empty vector; OMICSLINK™ CARF Lentiviral ORF expression clone]
Hasan, M.N., et al. (2023). Combining ASBT inhibitor and FGF15 treatments enhances therapeutic efficacy against cholangiopathy in female but not male Cyp2c70 KO mice. J Lipid Res doi: 10.1016/j.jlr.2023.100340 [OMICSLINK™ Fgf15 ORF expression clone; AAVPrime™ AAV particles expressing Fgf15]
Gannon, A-L., et al. (2022). A Novel Model Using AAV9-Cre to Knockout Adult Leydig Cell Gene Expression Reveals a Physiological Role of Glucocorticoid Receptor Signalling in Leydig Cell Function Int J Mol Sci doi: 10.3390/ijms232315015 [AAVPrime™ AAV particles expressing CRE and GFP]
Chen, X., et al. (2022). Pitx3 deficiency promotes age-dependent alterations in striatal medium spiny neurons. Front Aging Neurosci doi: 10.3389/fnagi.2022.960479 [AAV-hSyn1-eGFP]
Yang, B., et al. (2022). Evaluation of Early Biomarkers of Atherosclerosis Associated with Polychlorinated Biphenyl Exposure: An in Vitro and in Vivo Study. Environmental Health Perspectives doi: 10.1289/EHP9833 [AAVPrime™ AAV particles expressing HDAC7]
Xia, Y., et al. (2022). Neuronal C/EBPb/AEP pathway shortens life span via selective GABAnergic neuronal degeneration by FOXO repression. Science Advances doi: 10.1126/sciadv.abj8658 [AAVPrime™ AAV particles expressing FOXO1, shRNA targeting FOXO1]
Song, M-Y., et al. (2022). Sirt6 reprograms myofibers to oxidative type through CREB-dependent Sox6 suppression. Nature Communications doi: 10.1038/s41467-022-29472-5 [AAVPrime™ AAV particles expressing Sirt6, shRNA targeting Sirt6]
Lio, J., et al. (2022). C/EBPβ/AEP signaling couples atherosclerosis to the pathogenesis of Alzheimer’s disease. Molecular Psychiatry doi: 10.1038/s41380-022-01556-0 [AAVPrime™ AAV particles expressing shRNA targeting ApoE]
Toan, N.K., et al. (2021). Choline Acetyltransferase Induces the Functional Regeneration of the Salivary Gland in Aging SAMP1/Kl -/- Mice. International Journal of Molecular Sciences doi: 10.3390/ijms22010404 [AAVPrime™ AAV particles expressing shRNA targeting ChAT]
Zhang, W-B., et al. (2021). Activation of Nrf2 by miR-152 Inhibits Doxorubicin-InducedCardiotoxicity via Attenuation of Oxidative Stress, Inflammation,and Apoptosis. Oxidative Medicine and Cellular Longevity doi: 10.1155/2021/8860883 [AAVPrime™ AAV particles expressing mir-152 miRNA; AAVPrime™ AAV particles expressing shRNA targeting Nrf2]
Kang, S.S., et al. (2021). ApoE4 inhibition of VMAT2 in the locus coeruleus exacerbates Tau pathology in Alzheimer’s disease. Acta Neuropathologica doi: 10.1007/s00401-021-02315-1 [AAVPrime™ AAV particles expressing ApoE4]
Quan, D.N. and Shiloach, J. (2021). rAAV production and titration at the microscale for high- throughput screening. Human Gene Therapy doi: 10.1089/hum.2021.080 [AAVPrime™ AAV particles expressing EGFP]
Han, L., et al. (2020). Homocysteine-induced electrical remodeling via the mediation of IP3R1/Nav1.5 signaling pathway. Am J Transl Res. 2020; 12(7): 3822–3841 [AAVPrime™ AAV particles expressing shRNA targeting IP3R1]
Luo, Y., et al. (2020). Upregulation of circ_0000199 in circulating exosomes is associated with survival outcome in OSCC. Scientific Reports doi: 10.1038/s41598-020-70747-y [AAVPrime™ AAV particles expressing shRNA targeting circ_0000199]
Luo, Y., et al. (2020). A novel epigenetic regulation of circFoxp1 on Foxp1 in colon cancer cells. Cell Death & Disease doi: 10.1038/s41419-020-03007-6 [AAVPrime™ AAV particles expressing shRNA targeting circ_FoxP1]
Millett, P.J., et al. (2020). Gene Editing to Impove Joint Function. United States Patent Applicaitons doi: 20200360454K [AAVPrime™ AAV particles expressing GFP]
Zhao, Y. and Ren, J. (2019). MODIFIED IMMUNE CELLS HAVING ENHANCED FUNCTION AND METHODS FOR SCREENING FOR SAME. United States Patent Application 20190345491 [AAVPrime™ AAV particles expressing DNMT3A]

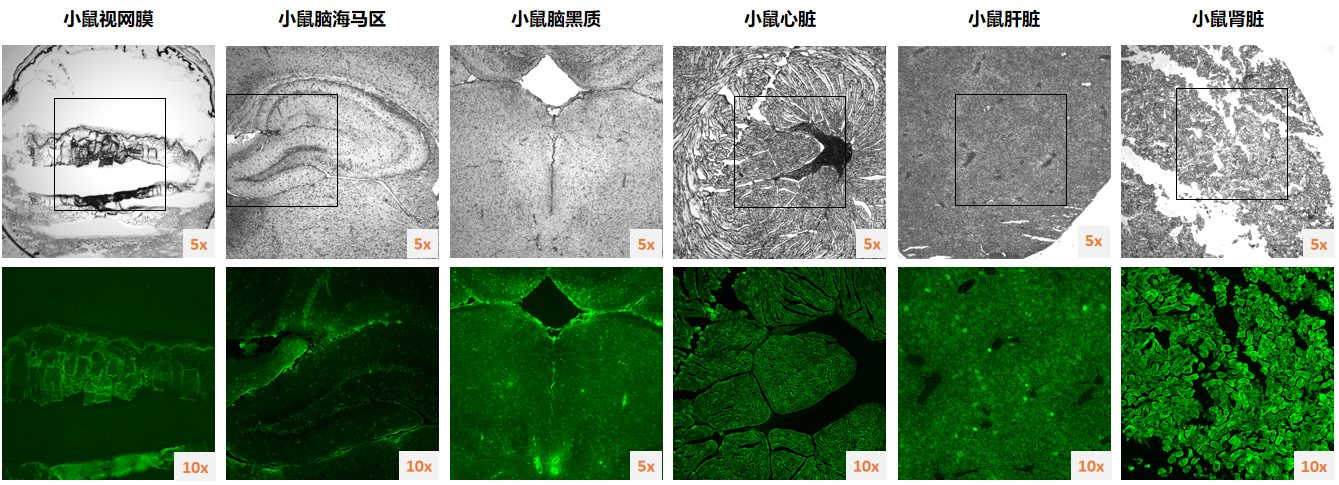
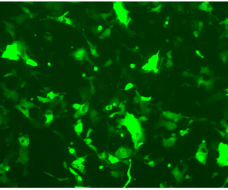
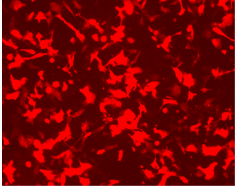
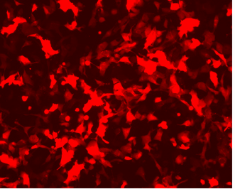
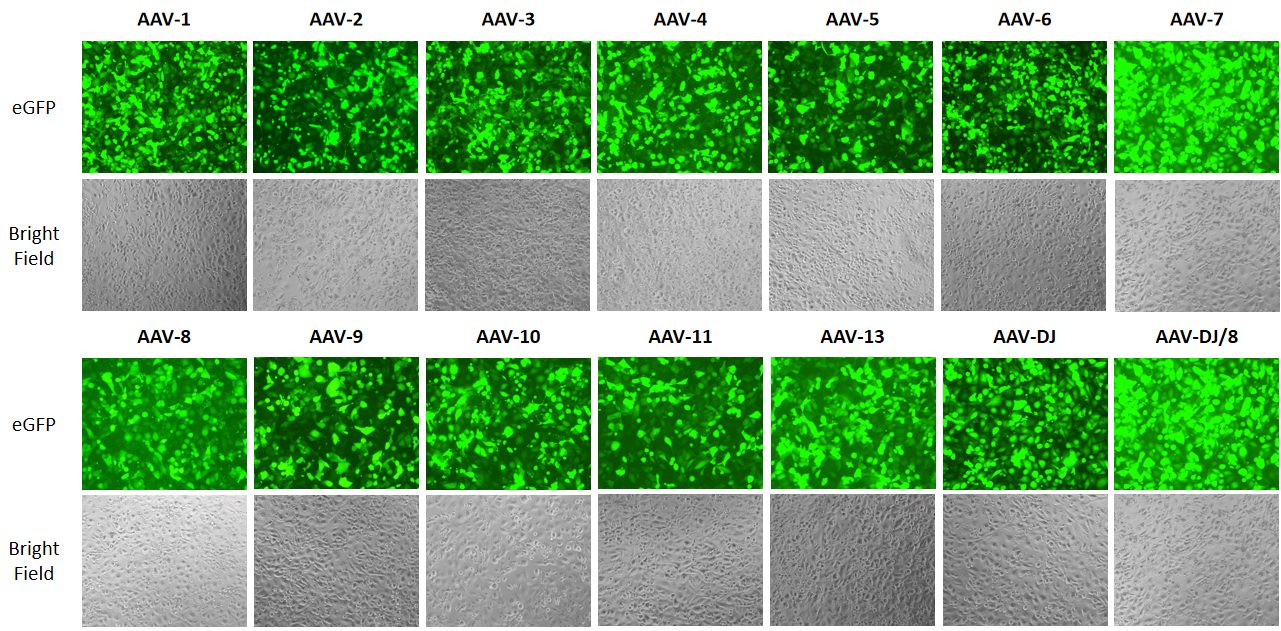
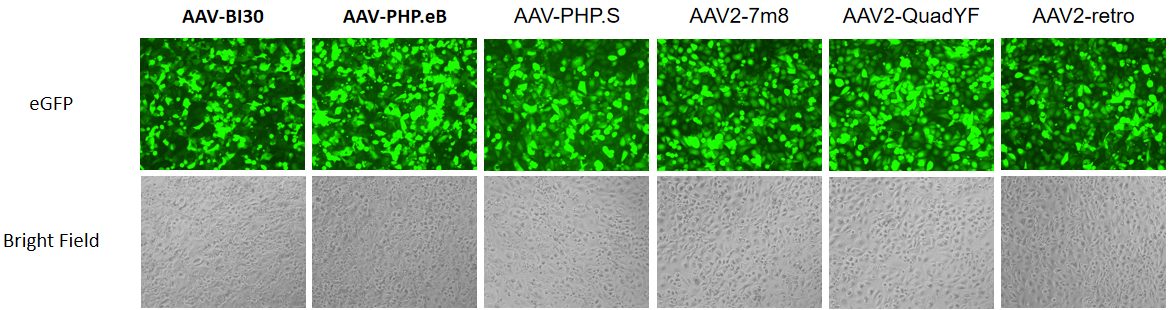
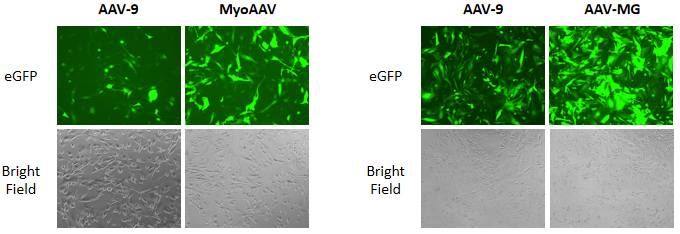
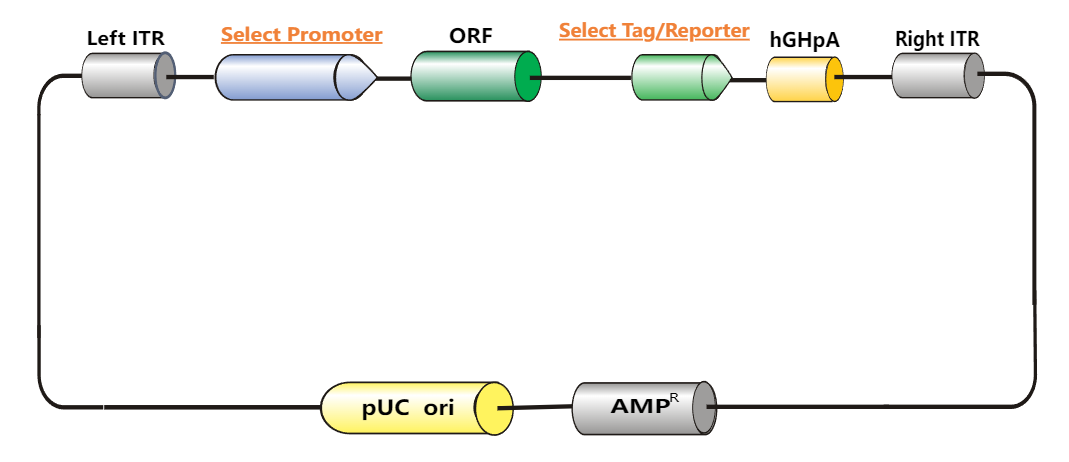




{kind=link}