
免疫系统通常不会攻击人的自身机体,当自身免疫性疾病发生时,则会产生自我反应的T细胞或循环抗体,去识别宿主蛋白(也称自身抗原)。已有超过100种自身免疫疾病被发现,因此急需能够高灵敏度检测大量自身抗体的工具。
OmicsArray™ 系统性自身免疫疾病相关的自身抗体检测服务使用高灵敏度的抗原微阵列芯片,能够高效地检测与许多自免疾病相关的自身抗体,包括类风湿关节炎、肌肉营养不良、系统性红斑狼疮和 I 型糖尿病。芯片中包含的 120 种抗原是根据已发表的研究结果选择的。高质量的纯化抗原被涂布在微芯片的三维表面上,以确保抗原保持其天然构象。每个阵列都包含内部 Ig 和抗 Ig 对照,用于归一化和量化。此外,在我们的微阵列中使用荧光二抗,可以在同一个阵列上检测两种类型(IgG、IgM 或 IgA)的抗体,从而最大限度地减少了所需的样品。
*PA001-V2和PA001-V3是PA001的两个版本,两个版本之间有19种抗原的差异,PA001-V3的特有抗原大多与炎性肌炎有关。
OmicsArray™ 自身抗体检测服务(抗原芯片)
自身抗体与多种自身免疫性疾病和免疫系统失调有关。自身抗体检测服务主要用于自身抗体的检测,用抗原芯片分析针对一系列自身抗原的自身抗体图谱,可对自身免疫性疾病、过敏、肿瘤反应、接种疫苗和感染、器官移植等的免疫反应进行大规模分析,为疾病预警和诊断、治疗方法选择、疗效评估、预后分析等科学研究提供依据,也是发现具有临床价值的抗体标记、研究疾病发病机制的有力工具。 Genecopoeia提供用于转化医学研究的,基于抗原芯片的自身抗体检测服务。OmicsArray™自身抗体检测芯片能用于以下研究,包括:
-
- 各种自身免疫性疾病,如:系统性红斑狼疮、类风湿性关节炎、糖尿病、混合结缔组织病、Sjøgren综合症、硬皮病、多肌炎、皮肌炎等;
- 过敏性疾病;
- 传染性疾病;
- 脑神经失调或神经障碍;
- 病毒疫苗的开发及疫苗接种后对自身免疫反应的监测,如SARS-CoV-2、其他冠状病毒株、流感病毒和呼吸道合胞病毒(RSV)等。
- 癌症生物标志物和免疫调节治疗毒性监测;
- 器官移植效果评估;
- 药物筛选和临床试验。
产品优势
- 高多重性: 与传统的ELISA法每孔检测1个抗体相比,自身抗原阵列每孔可检测>120个自身抗体;
- 高通量: 每张玻片可同时处理16个样品,多张玻片可并行操作;
- 高灵活性: 结合不同抗原(蛋白质、多肽、核酸等),检测不同类型和亚型的抗体;
- 小样本量: 5ul血清或其他体液即可满足;
- 成本低: 低于ELISA试剂盒1/10的价格;
- 高灵敏度: 结果的动态范围在1-65000;
- 半定量: 基于标准曲线的相对定量。
技术简介
- 什么是自身抗体/抗原? 在自身免疫性疾病发生、发展的过程中,引起自身免疫应答的自身组织成分被称为自身抗原,包括隐藏的自身抗原(在胚胎期从未与自身淋巴细胞接触过,机体不能识别为自身物质)和修饰的自身抗原(在感染、药物、烧伤、电离辐射等因素影响下,自身组织的构象发生改变,成为自身抗原)。机体产生自身抗原时,由于免疫系统将其识别为异己部分,会产生对应的自身抗体进行抵抗。
- 什么是抗原芯片? 自身抗原芯片,是指以蛋白质分子作为配基,将其有序地固定在固相载体的表面形成微阵列,用标记了荧光的蛋白质或其他它分子与之作用,洗去未结合的成分,经荧光扫描等检测方式测定芯片上各点的荧光强度,来分析蛋白之间或蛋白与其它分子之间的相互作用关系。
- 为什么要做这个服务? 这些自身抗原或自身抗体在疾病表型显现的前期往往已经产生,并且表达量通常随着病情的演化呈现出一定的趋势,是一类重要的疾病生物标志物。越来越多科研人员将目光定格在自身抗体相关标志物的研究上,力求在自身抗体层面寻找生物标志物,建立诊断、预后、药效评估的多参数模型。 为了满足疾病诊断,预后判断等多方面的广泛而长期的需求,疾病生物标志物的发现研究不可或缺。基于疾病相关自身抗体的标志物因在多种疾病中广泛存在,且可通过血液样本而方便地实现检测而备受关注。
服务流程
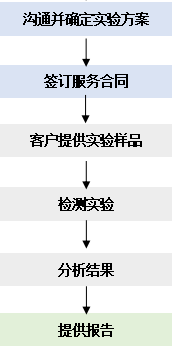
点击下载 服务指南 
样本要求
1. 适用于自身抗原芯片检测的常见样品 血浆、血清、淋巴、尿液、间质液、渗出液、细胞溶解液、分泌液等体液均可用于自身抗原芯片检测分析。
2. 样品的收集方法 a. 血清:采用不含抗凝剂的采血管采集全血,4℃放置30-45min后,4℃ 3000rpm离心10min,取上清,存于-80℃冰箱;b. 血浆:采用抗凝剂的采血管采集全血,4℃放置30-45min后,4℃ 3000rpm离心10min,取上清,存于-80℃冰箱;c. 尿液:收集不添加稳定剂的尿液样本后,4℃ 10000rpm离心1min或5000rpm离心2min,取上清,在干冰/甲醇浴中快速冷冻,储存在-80℃备用。
3. 样本量 为保证实验的顺利进行,体液样本的样本量请≥50 ul。
4. 样本寄送要求 a. 快递发出后请于当天将快递单号或专业化冷链物流托运单号及时告知乙方; b. 收货后乙方会对样品到达状态拍照,若样本送达状态不佳,可能需重新寄送; c. 体液样本请优先保存于干冰中寄出,干冰用量需足够以确保寄到时仍有干冰剩余; d. 不能使用干冰的情况下请使用冰袋,冰袋数量需足够以确保寄到使样本温度低于4℃。
数据分析
GeneCopoeia的抗原芯片检测服务含简要数据分析。阵列扫描后,使用GenePix®7.0软件收集和分析原始数据。数据分析包括数据归一化、差异分析、聚类分析,以及按变化水平排列的每个抗原的热图。我们的分析按照以下步骤:
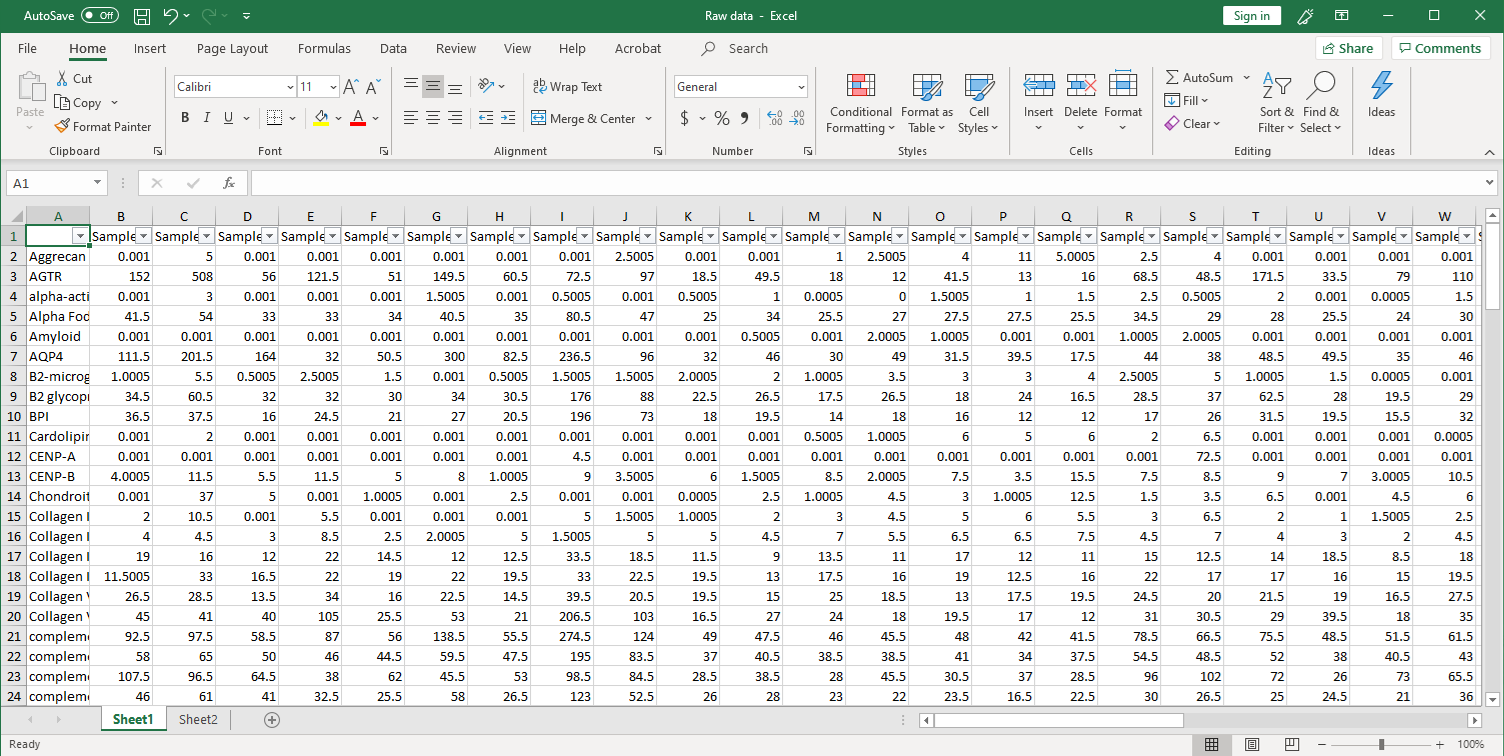
1. 收集原始数据。扫描后,在Excel文件中收集原始信号强度,如下所示。
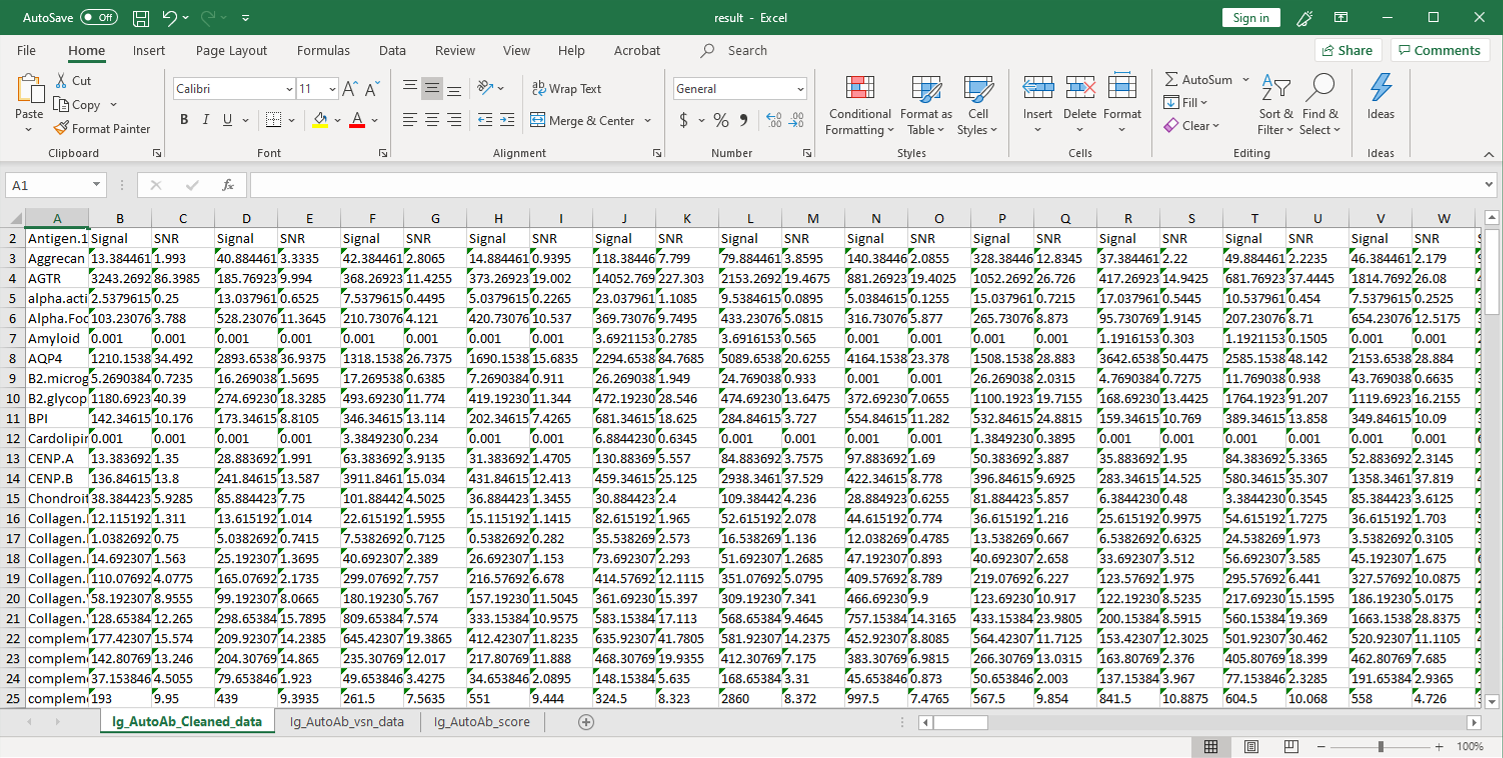
2. 然后,将原始数据用参照点的值进行归一化,并将净信号强度(Net Signal Intensity, NSI)值以及信噪比(signal-to-noise ratios, SNRs)列在一个Excel文件中。
3. 标准分析的最后一步是确定“抗体评分”,即样品中给定抗体的相对富集程度。抗体评分以数字形式展示,并显示在热图中。得分越高(在热图上向红色移动)表明抗体-抗原相互作用越强,如下例所示:

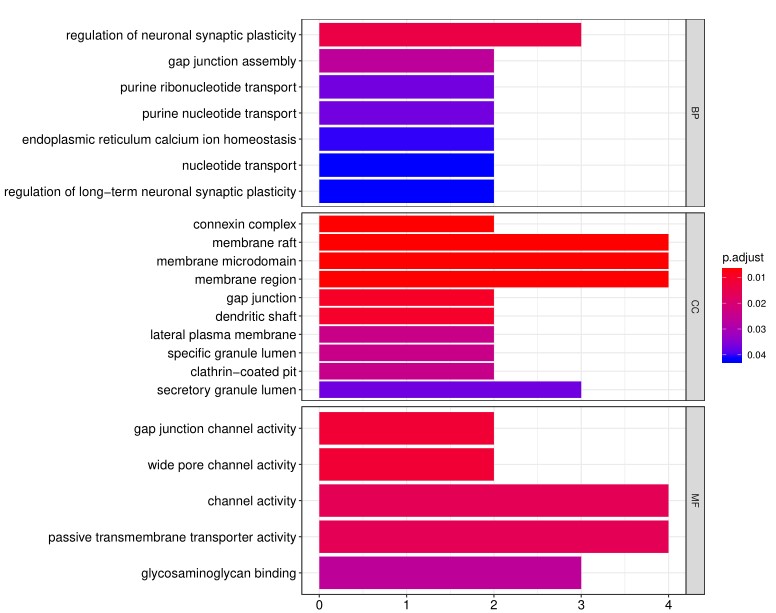
4. 除标准数据分析包外,客户还可以选择定制数据分析服务。其中一项服务是使用基因本体( Gene Ontology, GO)分析对阵列上的阳性抗原进行分类,该分析基于已知的生物功能对基因和蛋白质进行分类。分子功能GO分析的读数示例如下所示:
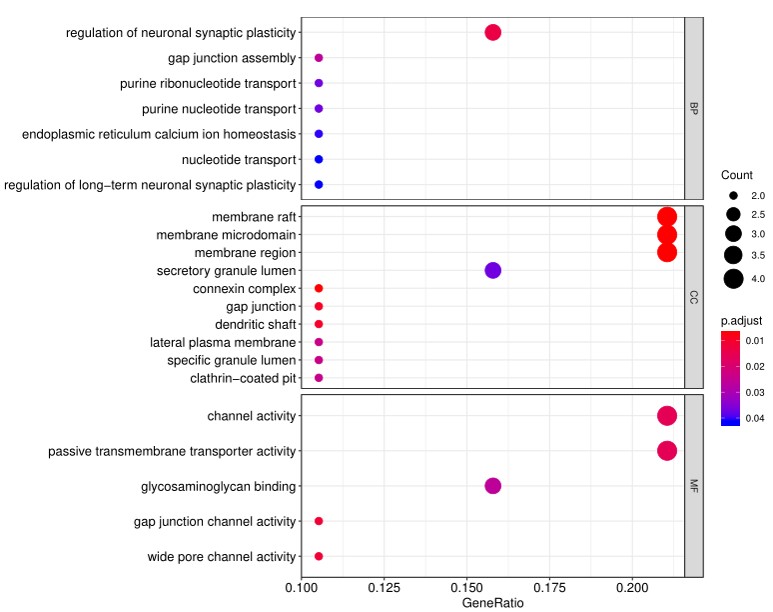
此外,生物过程的GO分析如下示例所示:
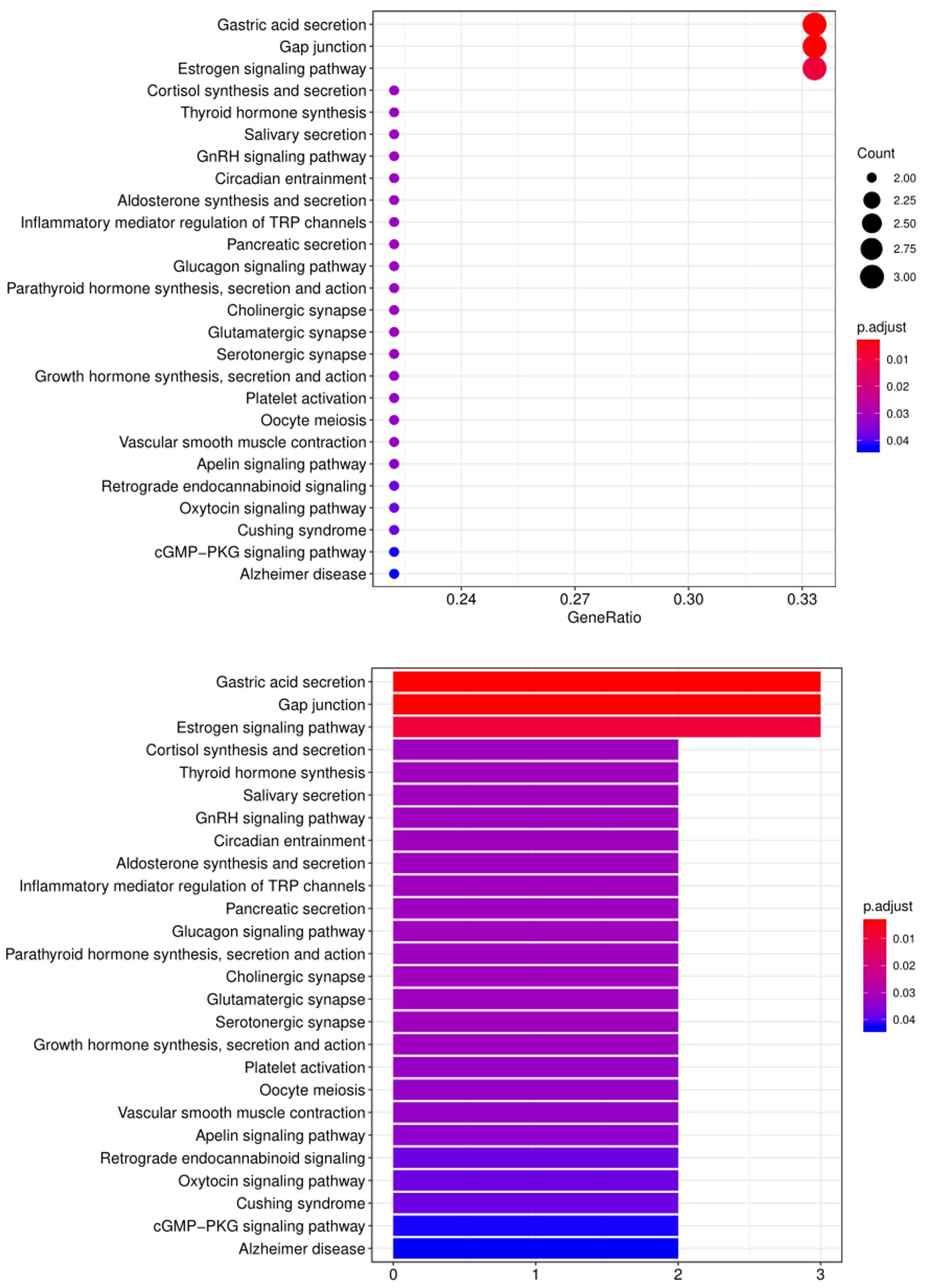
5. 另一个广泛用于生物信息分析的工具是KEGG (Kyoto Encyclopedia for Genes and Genomes) 通路分析,我们使用该工具根据已定义的生物通路对阳性抗体-抗原相互作用进行分组,如下所示:
报告形式
服务的实验结果将通过邮件形式提供相应的实验数据资料及报告文件。包括:
- 蛋白芯片扫描图;
- 杂交信号强度值;
- 差异分析;
- 聚类分析等。
FAQs
请在实验之前与售前进行充分沟通,说明实验目的,样本情况,以明确我方是否能满足您的需求以及确定实验方案。
抗原芯片上的抗原,全部是来自于文献报道的自身抗原。对于每种不同类型的抗原芯片,我们已展示相关抗原20个,在您确定 签署相应服务协议后,我们会给您提供全部的抗原及相关信息,在您使用自身抗原芯片的检测结果发表文章之前,请勿对第三方展示此信息。
- 由于每张芯片上有16个block,其中一个用来做PBS对照,剩下15个用来孵育样本。所以客户所送样本的总数量最好为15的倍数,这样可以充分保证芯片的使用效率,避免让客户分担相应的损失成本。
- 理论上每个不同的检测组最低需要3个样本才能满足统计学上重复性的要求。在此基础上,样本越多,其结果对检测组的代表性越高。为了保证结果的可靠性,推荐不同检测组样本量≥30。
抗原芯片采用荧光检测,其灵敏度要比ELISA(比色法)、Western(化学发光法)更加灵敏和稳定。片内和片间的技术重复相关系数R2均可以达到0.9以上。动态检测范围在1-65000。动态范围越大,说明所能检测的信号层次就越多。
如果您需要定制芯片,我们先需要与您就以下问题进行充分沟通:
- 抗原的知识产权情况(是来自于已公开发表的论文还是已受专利保护等),我们仅能提供不涉及专利保护的抗原定制检测;
- 抗原的基本信息(是全蛋白还是肽段,分子量,结构域,是否为膜蛋白等);
- 您是否能提供满足定制芯片要求的抗原;
- 若需要我方提供待检抗原,相应费用及实验周期需另外核算。
客户可以选择是否需进行预实验,若进行预实验,客户可提供15例样本(需符合样本要求),若您后续继续签署服务,且预实验的数据可用,再记入正常样品收费。
a. 这个差异也许与检测手段不同相关,芯片用的蛋白为非变性蛋白,和WB有所差别;芯片的杂交体系为抗原捕获抗体,与WB的抗原转膜后与抗体孵育也稍有差别,因此也许会造成部分差异; b. 需要客户提供WB验证的结果图,以便我们更好的分析分析客户验证的指标在芯片上的图像和数据情况;
目前应用自身抗原芯片发表的研究类文章已经非常多。我们的自身抗原芯片已在全世界近百家医疗科研和医药企业得到广泛应用,已独立或和用户合作发表了几十篇论文。其中高影响因子的论文含数十篇。蛋白芯片技术可以极大提高科学研究的效率,更能够获得较为全面的数据,以在研究中得到创新的结果和结论,是科学论文发表的加分项。
2024年
- Li, Q., et al. (2024). Phenotypic and Immunological Characterization of Patients with Activated PI3Kδ Syndrome 1 Presenting with Autoimmunity. J Clin Immunol. 44(4):102.
- Preeti S. Chauhan, et al. (2024). Dietary docosahexaenoic acid supplementation inhibits acute pulmonary transcriptional and autoantibody responses to a single crystalline silica exposure in lupus-prone mice. Front Immunol. 15:1275265.
2023年
- Son, K., et al. (2023). Circulating anti-nuclear autoantibodies in COVID-19 survivors predict long COVID symptoms. Eur Respir J. 61(1):2200970.
2022年
- Heine, L.K., et al. (2022). Comparative effects of human-equivalent low, moderate, and high dose oral prednisone intake on autoimmunity and glucocorticoid-related toxicity in a murine model of environmental-triggered lupus. Front Immunol. 13:972108.
- Khassawneh, B., et al. (2022). Autoantibody profile in sarcoidosis, analysis from the GRADS sarcoidosis cohort. PLoS One. 17(10):e0274381.
- Ning, W., et al. (2022). Staphylococcus aureus peptidoglycan (PGN) induces pathogenic autoantibody production via autoreactive B cell receptor clonal selection, implications in systemic lupus erythematosus. J Autoimmun. 131:102860.
- Salter, B., et al. (2022). Airway autoantibodies are determinants of asthma severity. Eur Respir J. 60(6):2200442.
- Elias, S., et al. (2022). CXCR4+ Treg cells control serum IgM levels and natural IgM autoantibody production by B-1 cells in the bone marrow. J Exp Med. 219(7):e20220047.
- von Itzstein, M.S., et al. Association between Antibiotic Exposure and Systemic Immune Parameters in Cancer Patients Receiving Checkpoint Inhibitor Therapy. Cancers (Basel). 14:1327.
- Seeley-Fallen MK, Lazzaro M, et al. (2022). Non-Muscle Myosin II Is Essential for the Negative Regulation of B-Cell Receptor Signaling and B-Cell Activation. Front Immunol. 13:842605.
-
Avalos A, Tietsort JT, et al. (2022). Hem-1 regulates protective humoral immunity and limits autoantibody production in a B cell-specific manner. JCI Insight. 7(9):e153597.
-
Labombarde JG, Pillai MR, et al. (2022). Induction of broadly reactive influenza antibodies increases susceptibility to autoimmunity. Cell Rep. 38(10):110482.
- Boustani K, Ghai P, et al. (2022). Autoantibodies are present in the bronchoalveolar lavage but not circulation in patients with fibrotic interstitial lung disease. ERJ Open Res. 8(1):00481-202.
-
Ghosh N, Postow M, et al. (2022). Lower baseline autoantibody levels are associated with immune-related adverse events from immune checkpoint inhibition. J Immunother Cancer. 10(1):e004008.
- Rojas M, Ramírez-Santana C, et al. (2022). New insights into the taxonomy of autoimmune diseases based on polyautoimmunity. J Autoimmun.126:102780.
- Cheng D, Luo Z, et al. (2022). Elevated Cerebrospinal Fluid Anti-CD4 Autoantibody Levels in HIV Associate with Neuroinflammation. Microbiol Spectr. 10(1):e0197521.
2021年
- Gonugunta AS, von Itzstein MS, et al. (2021). Humoral and cellular correlates of a novel immune-related adverse event and its treatment. J Immunother Cancer. 9(12):e003585.
- Chiang K, Largent AD, et al. (2021). Cutting Edge: A Threshold of B Cell Costimulatory Signals Is Required for Spontaneous Germinal Center Formation in Autoimmunity. J Immunol. 207(9):2217-2222.
- Hirsiger JR, Tamborrini G, et al. (2021). Chronic inflammation and extracellular matrix-specific autoimmunity following inadvertent periarticular influenza vaccination. J Autoimmun. 124:102714.
- Kiripolsky J, Kasperek EM, et al. (2021). Immune-Intrinsic Myd88 Directs the Production of Antibodies With Specificity for Extracellular Matrix Components in Primary Sjögren’s Syndrome. Front Immunol. 12:692216. doi: 10.3389/fimmu.2021.692216.
- Jacobs HM, Arkatkar T, et al. (2021). TACI haploinsufficiency protects against BAFF-driven humoral autoimmunity in mice. Eur J Immunol. 51(9):2225-2236.
- Cass SP, Dvorkin-Gheva A, et al. (2021). Differential expression of sputum and serum autoantibodies in patients with chronic obstructive pulmonary disease. Am J Physiol Lung Cell Mol Physiol. 320(6):L1169-L1182.
- Pestka JJ, Akbari P, et al. (2021). Omega-3 Polyunsaturated Fatty Acid Intervention Against Established Autoimmunity in a Murine Model of Toxicant-Triggered Lupus. Front Immunol. 12:653464.
- Wilson A, Velasco CA, et al. (2021). Mine-site derived particulate matter exposure exacerbates neurological and pulmonary inflammatory outcomes in an autoimmune mouse model. J Toxicol Environ Health A. 84(12):503-517.
- Kiripolsky J, Kasperek EM, et al. (2021). Tissue-specific activation of Myd88-dependent pathways governs disease severity in primary Sjögren’s syndrome. J Autoimmun. 118:102608.
- Rai P, Janardhan KS, et al. (2021). IRGM1 links mitochondrial quality control to autoimmunity. Nat Immunol. 2021 Mar;22(3):312-32.
- Garimella MG, He C, et al. (2021). The B cell response to both protein and nucleic acid antigens displayed on apoptotic cells are dependent on endosomal pattern recognition receptors. J Autoimmun. 117:102582.
- Luo Z, Alekseyenko AV, et al. (2021). Rigorous Plasma Microbiome Analysis Method Enables Disease Association Discovery in Clinic. Front Microbiol. 11:613268.
2020年
- Khan, S., et al. Late-Onset Immunotherapy Toxicity and Delayed Autoantibody Changes: Checkpoint Inhibitor-Induced Raynaud’s-Like Phenomenon. Oncologist. 25: e753.
- Fuchs PS, Lötscher J, et al. (2020). Co-Occurrence of ANCA-Associated Vasculitis and Sjögren’s Syndrome in a Patient With Acromegaly: A Case Report and Retrospective Single-Center Review of Acromegaly Patients. Front Immunol. 11:613130.
- Vo HTM, Duong V, et al. (2020). Autoantibody Profiling in Plasma of Dengue Virus-Infected Individuals. Pathogens. 9(12):1060.
- Turner JA, Stephen-Victor E, et al. (2020). Regulatory T Cell-Derived TGF-β1 Controls Multiple Checkpoints Governing Allergy and Autoimmunity. Immunity. 53(6):1331-1332.
- Raj P, Song R, et al. (2020). Deep sequencing reveals a DAP1 regulatory haplotype that potentiates autoimmunity in systemic lupus erythematosus. Genome Biol. 21(1):281.
- Lammert C, Zhu C, et al. (2020). Exploratory Study of Autoantibody Profiling in Drug-Induced Liver Injury with an Autoimmune Phenotype. Hepatol Commun. 4(11):1651-1663.
- Turner JA, Stephen-Victor E, et al. (2020). Regulatory T Cell-Derived TGF-β1 Controls Multiple Checkpoints Governing Allergy and Autoimmunity. Immunity. 53(6):1202-1214.e6.
- Mayer CT, Nieke JP, et al. (2020). An apoptosis-dependent checkpoint for autoimmunity in memory B and plasma cells. Proc Natl Acad Sci U S A. 117(40):24957-24963.
- Rajasinghe LD, Li QZ, et al. (2020). Omega-3 fatty acid intake suppresses induction of diverse autoantibody repertoire by crystalline silica in lupus-prone mice. J Autoimmunity. 53(7):415-433.
- Perez-Diez A, Wong CS, et al. (2020). Prevalence and pathogenicity of autoantibodies in patients with idiopathic CD4 lymphopenia. J Clin Invest. 130(10):5326-5337.
- Tanaka S, Ise W, et al. (2020). Tet2 and Tet3 in B cells are required to repress CD86 and prevent autoimmunity. Nat Immunol. 21(8): 950-961.
- Mohammed S, Vineetha NS, et al. (2020). Regulatory role of SphK1 in TLR7/9-dependent type I interferon response and autoimmunity. FASEB J. 34(3):4329-4347.
2019年
- Chen Y, Yu M, et al. (2019). CXCR5+PD-1+ follicular helper CD8 T cells control B cell tolerance. Nat Commun. 10(1):4415.
- Wang T, Marken J, et al. (2019). High TLR7 Expression Drives the Expansion of CD19+CD24hiCD38hi Transitional B Cells and Autoantibody Production in SLE Patients. Front Immunol. 10:1243.
- Mohammed S, Vineetha NS, et al. (2019). Examination of the role of sphingosine kinase 2 in a murine model of systemic lupus erythematosus. FASEB J. 33(6):7061-7071.
- Ogunrinde E, Zhou Z, et al. (2019). A Link Between Plasma Microbial Translocation, Microbiome, and Autoantibody Development in First-Degree Relatives of Systemic Lupus Erythematosus Patients. Arthritis Rheumatol. 71(11):1858-1868.
- Luo Z, Li M, et al. (2019). Systemic translocation of Staphylococcus drives autoantibody production in HIV disease. Microbiome.7(1):2.
- Mukherjee M, Thomas SR, et al. (2019). Sputum Antineutrophil Cytoplasmic Antibodies in Serum Antineutrophil Cytoplasmic Antibody-Negative Eosinophilic Granulomatosis with Polyangiitis. Am J Respir Crit Care Med. 199(2):158-170.
- Du SW, Arkatkar T, et al. (2019). Functional Characterization of CD11c+ Age-Associated B Cells as Memory B Cells. J Immunol. 203(11):2817-2826.
- Pouzolles M, Machado A, et al. (2019). Intrathymic adeno-associated virus gene transfer rapidly restores thymic function and long-term persistence of gene-corrected T cells. J Allergy Clin Immunol. 145(2):679-697.e5.
2018年
- Arkatkar T, Jacobs HM, et al. (2018). TACI deletion protects against progressive murine lupus nephritis induced by BAFF overexpression. Kidney Int. 94(4):728-740.
- Zhang Y, Burberry A, et al. (2018). The C9orf72-interacting protein Smcr8 is a negative regulator of autoimmunity and lysosomal exocytosis. Genes Dev. 32(13-14):929-943.
- Ottens K, Hinman RM, et al. (2018). Foxo3 Promotes Apoptosis of B Cell Receptor-Stimulated Immature B Cells, Thus Limiting the Window for Receptor Editing. J Immunol. 201(3):940-949.
- Palmer VL, Worth AN, et al. (2018). IL10 restrains autoreactive B cells in transgenic mice expressing inactive RAG1. Cell Immunol. 331:110-120.
- Preite S, Cannons JL, et al. (2018). Hyperactivated PI3Kδ promotes self and commensal reactivity at the expense of optimal humoral immunity. Nat Immunol. 19(9):986-1000.
- Chen X, Sun X, et al. (2018). An autoimmune disease variant of IgG1 modulates B cell activation and differentiation. Science. 362(6415):700-705.
较早之前
- Kramer, J.M., et al. Analysis of IgM antibody production and repertoire in a mouse model of Sjögren’s syndrome. J. Leukoc. Biol. 99: 321.
- Packard, T.A., et al. COPD is associated with production of autoantibodies to a broad spectrum of self-antigens, correlative with disease phenotype. Immunol. Res. 55: 48.



