
-RNA导向的精确、灵活的基因组编辑工具 |
||
 |
 |
 |
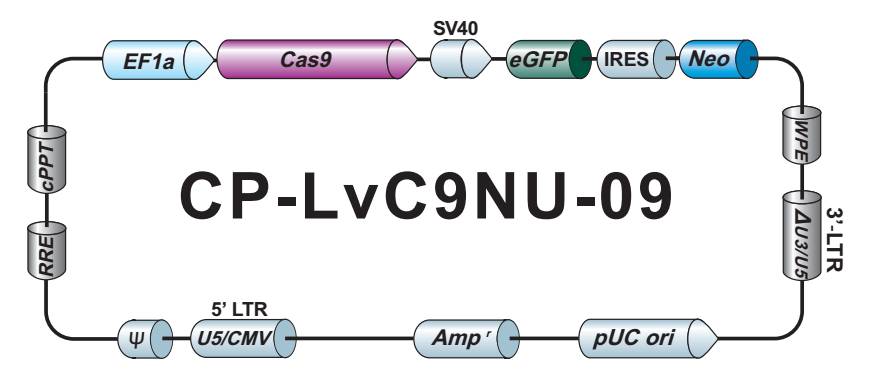
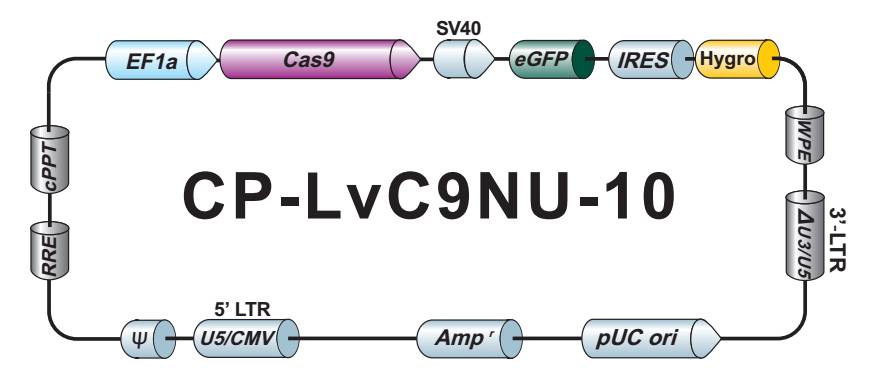
GeneCopoeia GeneHero™CRISPR-Cas9产品和服务为您的基因组编辑需求提供了完整、强大的解决方案。 产品和服务包括:
- 用于人、小鼠和大鼠的全基因组 sgRNA 克隆。 在我们的数据库中可搜索超过45,000个经过序列验证的、可用CRISPR 技术介导敲除的人、小鼠和大鼠基因。
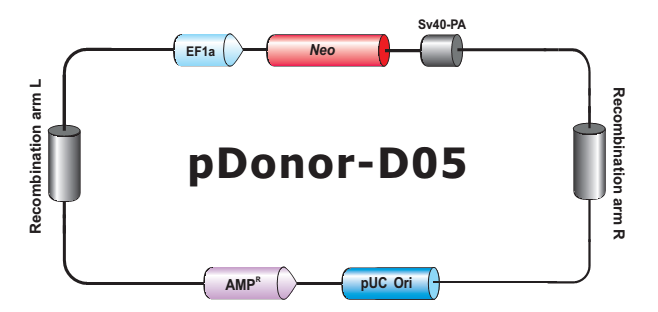
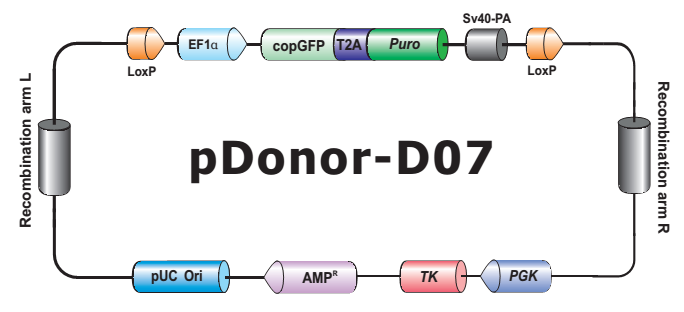
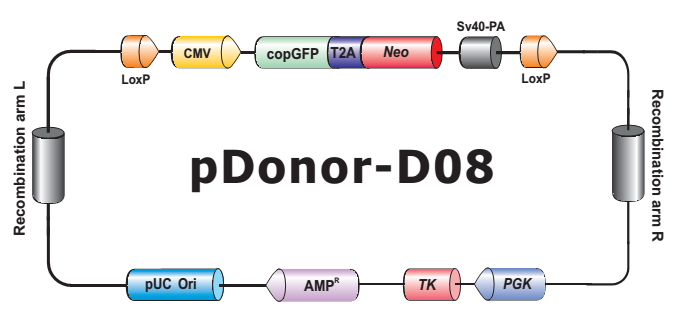
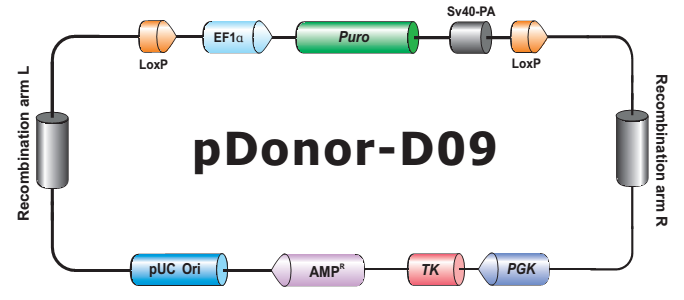
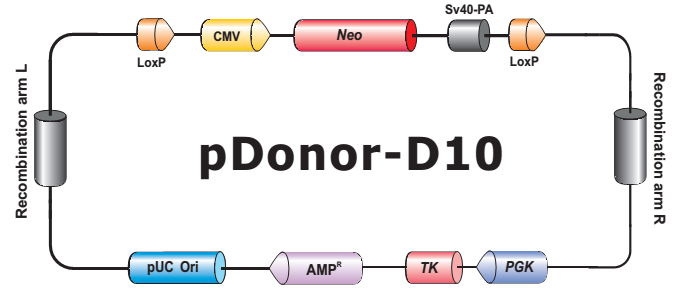
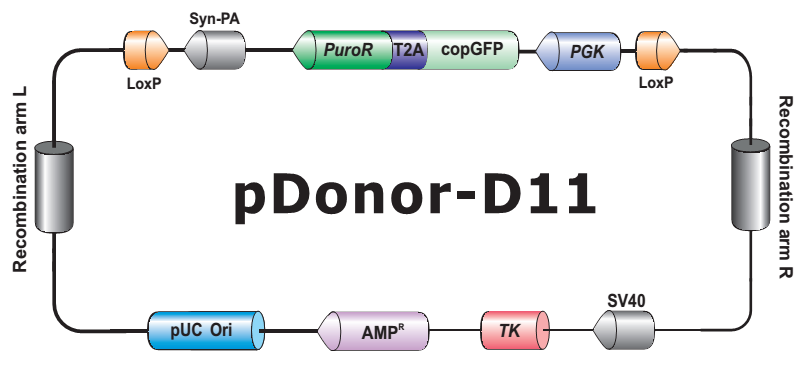
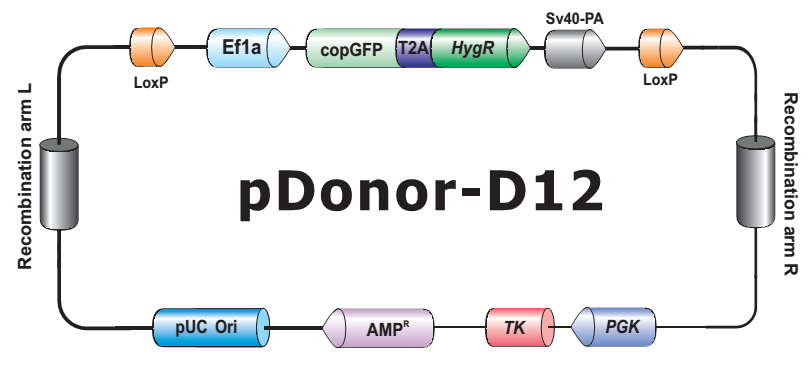
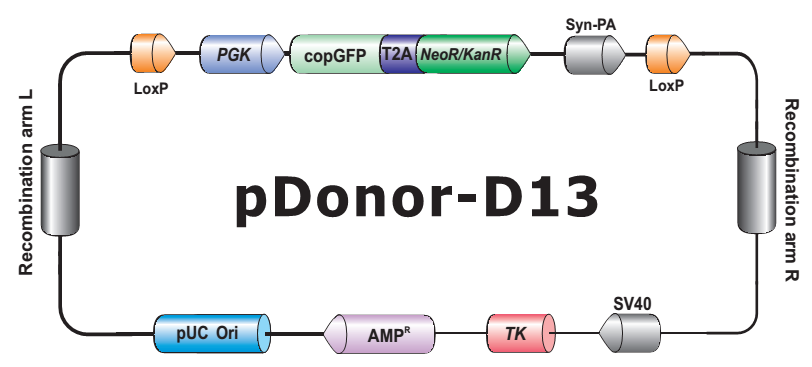
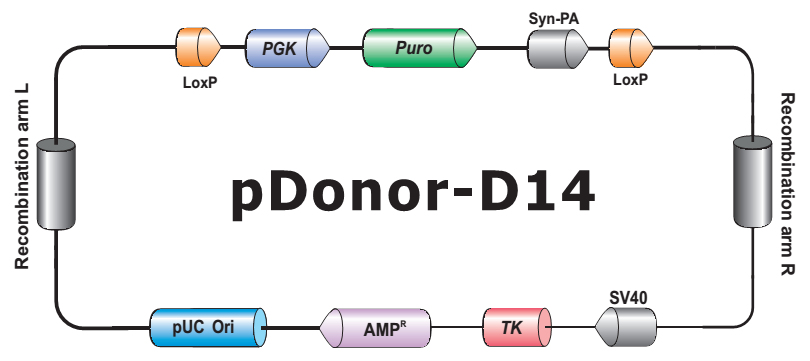
- HDR 供体克隆载体和定制的 HDR 供体克隆构建。可用于通过同源重组介导的 CRISPR 基因组编辑应用。
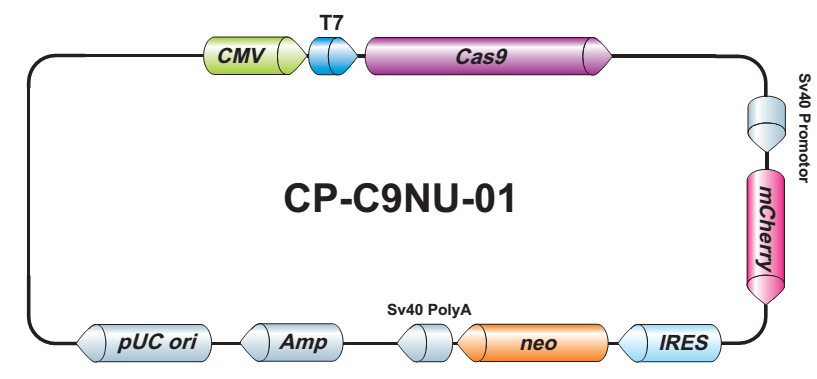
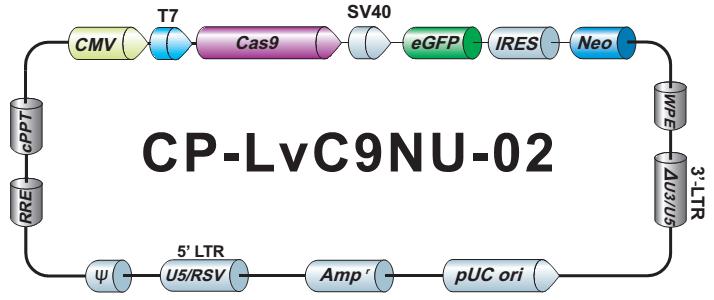
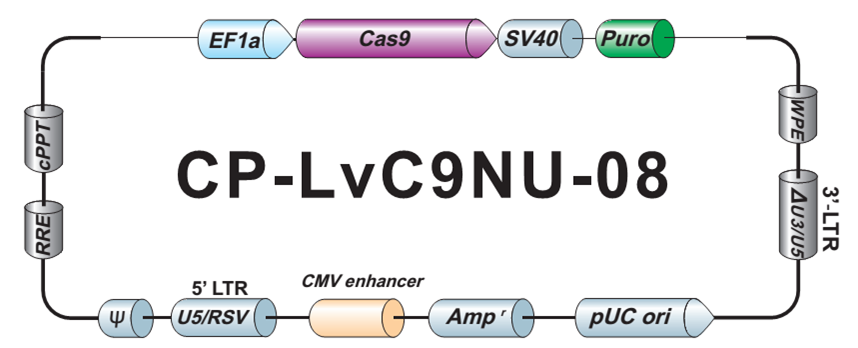
- Cas9稳定细胞系。预制 Cas9 稳定表达细胞系特别适用于sgRNA文库筛选和其他高通量 CRISPR-Cas9 应用。
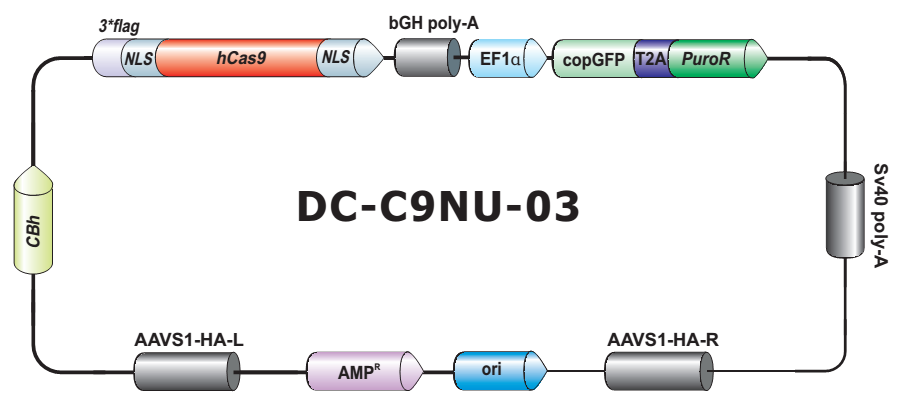
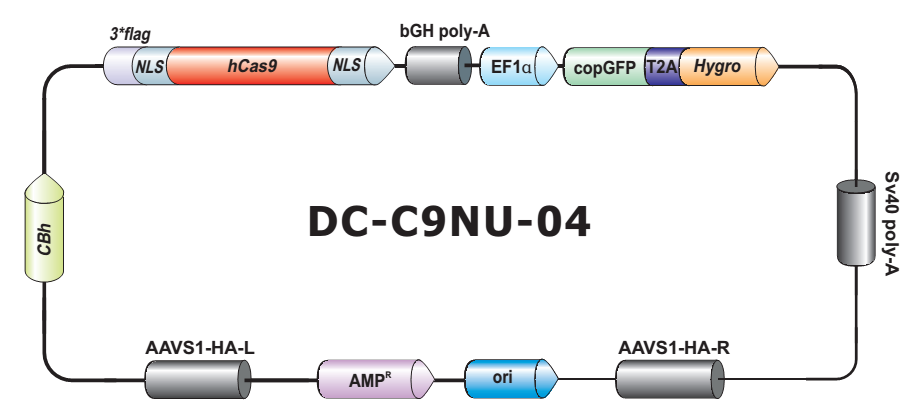
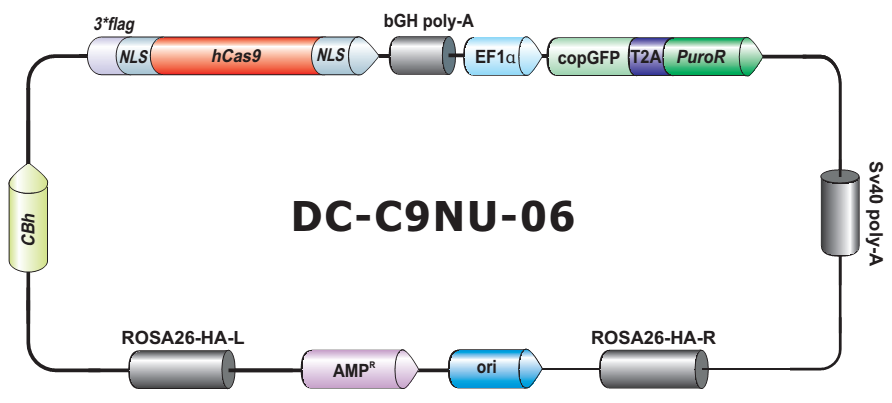
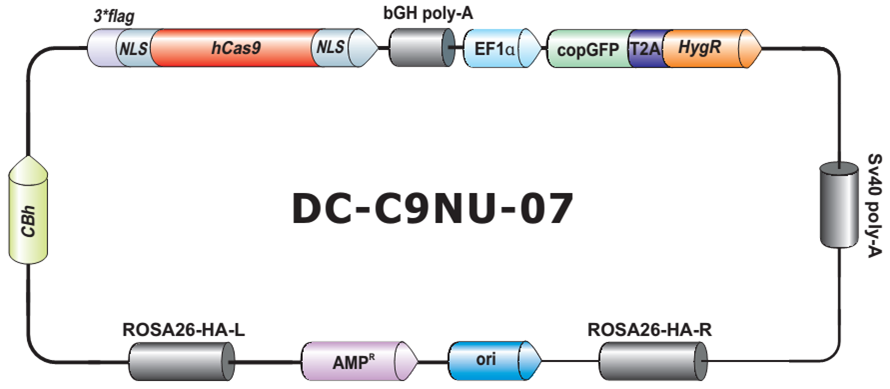
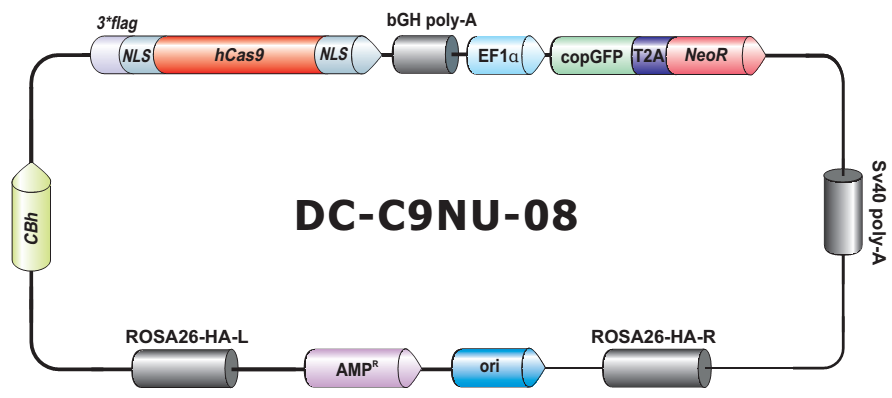
- Safe Harbor 基因敲入试剂盒及 ORF 敲入克隆。使用我们的人类 AAVS1 和小鼠 ROSA26Safe Harbor 基因敲入试剂盒可以轻松实现几乎任何DNA片段的成功敲入。人类及小鼠各有超过20,000条经过序列验证的ORF可用于转入基因的供体克隆设计。
- IndelCheck™ 插入缺失检测体系。IndelCheck™ CRISPR/TALEN 插入缺失检测体系是一套设计用于验证基因组编辑工具功能或检测编辑阳性细胞的完整体系,包括靶点 PCR 试剂盒、T7 核酸内切酶 I 检测试剂盒以及靶点 PCR 克隆试剂盒。另外,靶位点特异性PCR引物亦可选择。
- sgRNA 文库。我们提供7款预制的、通路特异性的 sgRNA 文库,每款都有慢病毒颗粒、转染级质粒 DNA 以及菌种等三种形式可供选择。客户也可以自行定制满足实验需求的 sgRNA 文库。
- VividFISH™染色体FISH计数探针。FISH是帮助您鉴定靶基因拷贝数的强大工具。在进行基因组编辑实验前使用VividFISH™探针来鉴定目的染色体拷贝数,可对后续筛选目的修饰阳性克隆有所帮助。
优势
- RNA导向的基因组DNA识别,无需考虑DNA甲基化
- 较ZFN和TALEN,有相同或更高的基因编辑效率
- 可同时编辑多个基因(多重靶向编辑)
- 设计简单快速,无需重复构建核酸内切酶
CRISPR工作原理
CRISPR-Cas系统(The clustered, regularly interspaced, short palindromic repeats-associated protein systems)是一个细菌及古细菌进化出来用以抵御病毒和质粒入侵的适应性机制。CRISPR-Cas系统的高效基因组编辑功能已被应用于多种生物,包括斑马鱼、小鼠、大鼠、秀丽隐杆线虫、植物及细菌。多个科研小组的研究都显示,与锌指核酸酶(ZFNs)和转录激活样效应核酸酶(Transcription activator-like effector nucleases, TALEN)相比较,CRISPR-Cas系统介导的基因组靶向实验在细胞或斑马鱼中具有相似甚至更高的效率。
CRISPR-Cas9体系的RNA-DNA识别机制为基因组工程研究提供了一项简便而强大的工具。该体系其中一个最重要的优势是Cas9蛋白可在多个不同的gRNA的引导下同时靶向多个基因组位点。
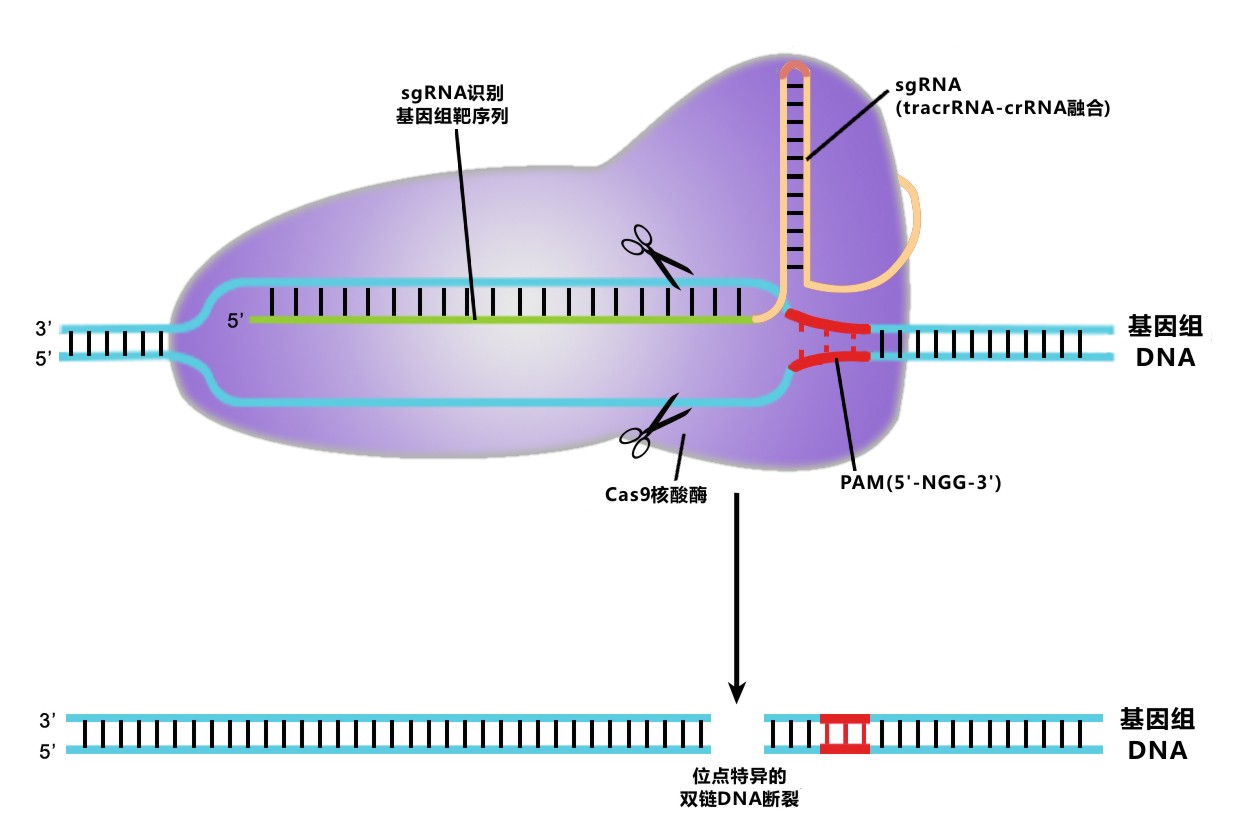
图 1. CRISPR/Cas9介导的基因组编辑原理图
在II型CRISPR系统中,CRISPR RNA(crRNA)与转录激活crRNA(Trans-activating crRNA, tracrRNA)退火形成的复合物能特异识别基因组序列,引导Cas9核酸内切酶在目的片段生成DNA双链断裂(double-strand breaks, DSBs)。 这个识别复合体可以通过融合crRNA与tracrRNA序列形成sgRNA(single-guided RNA)进行简化。基因组的靶序列中有长约20bp的片段与crRNA或sgRNA互补配对;靶序列末端的三核苷酸区域PAM(5’-NGG-3’)为Cas9识别位点,是实现剪切功能的关键。
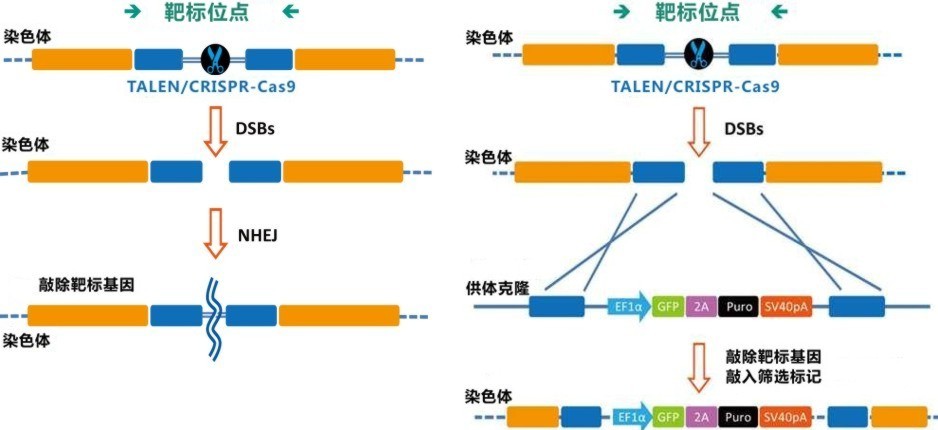
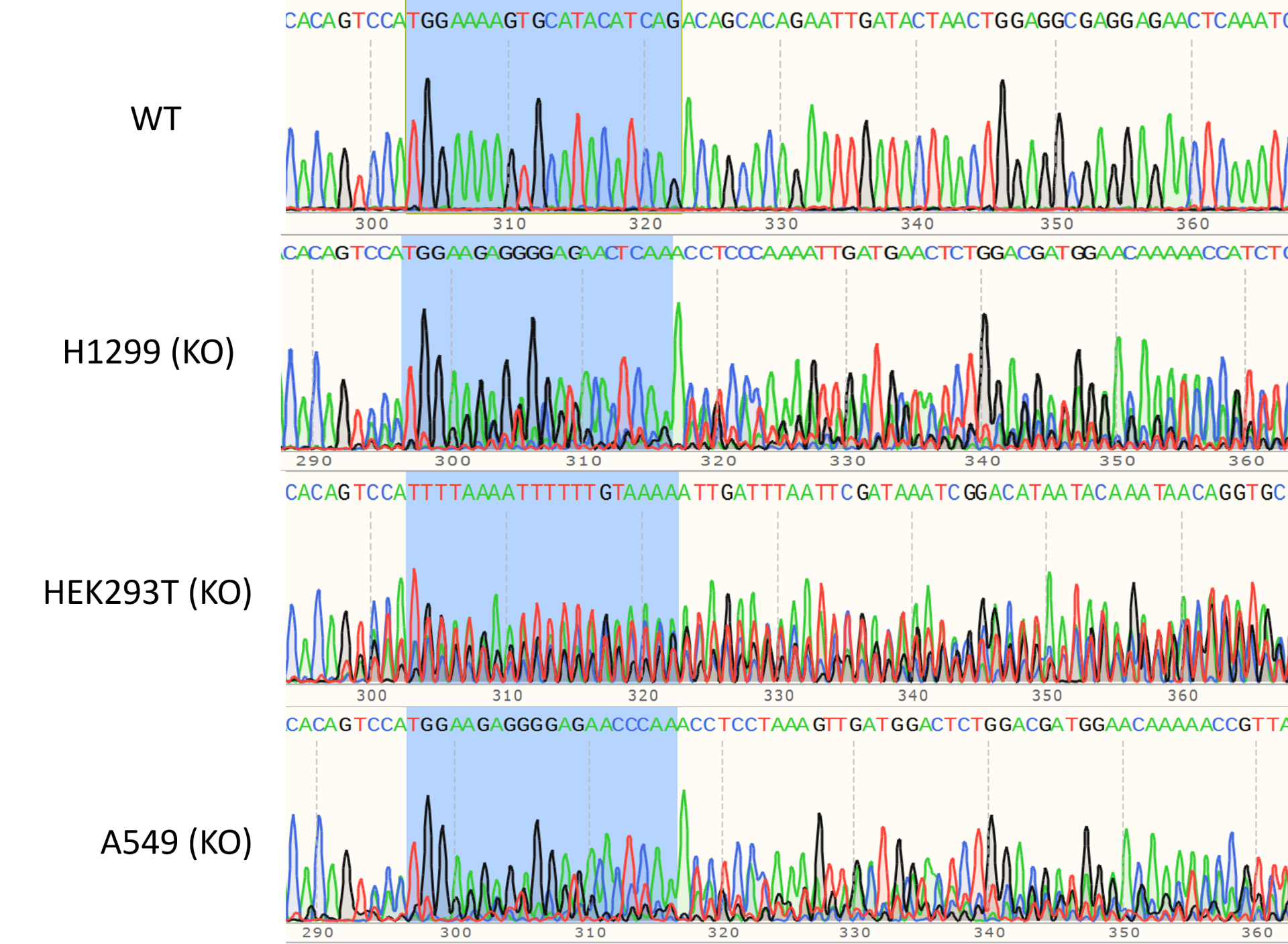
图2. CRISPR-Cas9介导的基因编辑。(左):sgRNA引导Cas9核酸酶作用于基因组,形成的DSBs被非同源末端连接(NHEJ)机制修复;(右):sgRNA引导Cas9核酸酶作用于基因组,形成DSBs,同源重组(HR)作用下,供体质粒上的目标基因及筛选标记(或其他遗传元件)在断裂处被整合进基因组。
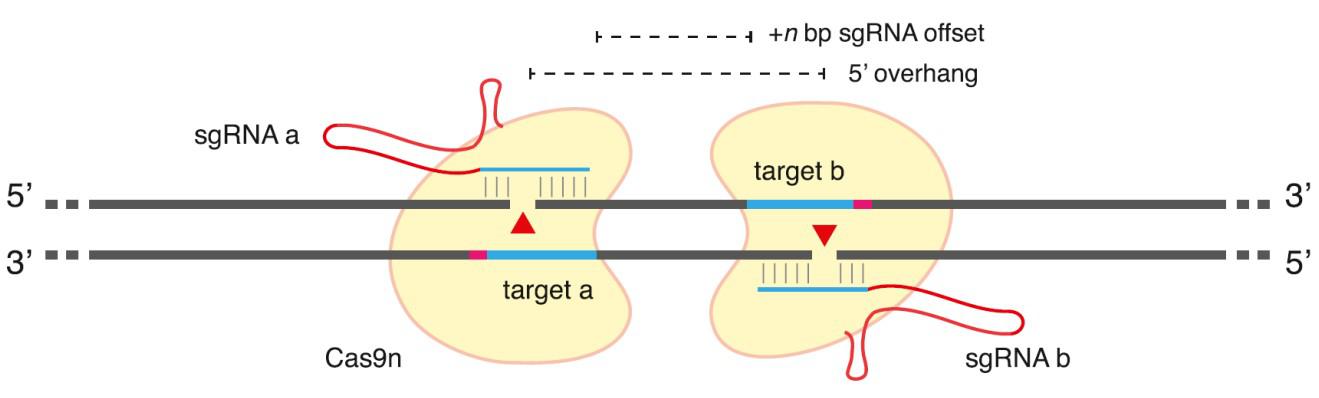
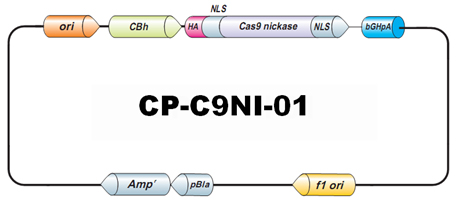
图3: 切口酶在结合链生产单链切口的原理图
TALEN 与 CRISPR-Cas9 体系对比
|
特性 |
TALEN |
CRISPR-Cas9 |
|---|---|---|
| 识别方式 |
蛋白质 – DNA |
RNA-DNA |
|
甲基化敏感性 |
敏感 |
不敏感 |
|
染色质结构敏感性 |
敏感 |
敏感 |
|
脱靶效应 |
较少观察到脱靶效应 |
潜在脱靶效应高于 TALEN 及ZFN |
|
多靶点操作 |
极少用 |
可用 |
基因组编辑应用
转录激活样效应因子(TAL Effectors)和CRISPR-Cas9可高效且精确地改造细胞系及模式动物的基因组。具体的基因组编辑应用可见下表。
| 基因组修饰 | 描述 | 基因组编辑工具 | 供体DNA |
|---|---|---|---|
| 内源基因标记 | 利用上融合标签(如荧光素酶、GFP等)跟踪内源性启动子活性或内源性蛋白的表达与定位。 | TALEN 或CRISPR | 必需 |
| 内源基因突变 | 在内源基因序列中引入点突变 | TALEN 或CRISPR | 必需 |
| 内源基因敲除 | 引入缺失或插入(如:筛选标记)敲除内源基因 | TALEN 或CRISPR | 若需建立敲除细胞系,则强烈建议使用。 |
| 内源基因表达激活 | 靶向激活内源基因表达 | TALE-TF或 CRISPR-TF | 不适用 |
| 内源基因表达抑制 | 靶向抑制内源基因表达 | TALE-R 或CRISPR-R | 不适用 |
| Safe harbor 位点敲入 | 敲入外源ORF或其他遗传因子到人或小鼠基因组的safe harbor位点 | TALEN或CRISPR | 必需 |
参考文献
- Horvath P, Barrangou R (January 2010). “CRISPR/Cas, the immune system of bacteria and archaea”. Science 327 (5962): 167–70.
- Marraffini LA, Sontheimer EJ (February 2010). “CRISPR interference: RNA-directed adaptive immunity in bacteria and archaea”. Nat Rev Genet 11 (3): 181–190.
- Hale CR, Zhao P, Olson S, et al. (November 2009). “RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex”. Cell 139 (5): 945–56.
- van der Oost J, Brouns SJ (November 2009). “RNAi: prokaryotes get in on the act”. Cell 139 (5): 863–5. doi:10.1016/j.cell.2009.11.018.
- Hale CR, Zhao P, Olson S, et al. (November 2009). “RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex”. Cell 139 (5): 945–56.
- Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J.A., and Charpentie E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptiv bacterial immunity. Science 337, 816–821.
- Jiang, W., Bikard, D., Cox, D., Zhang, F., and Marraffini, L.A. (2013). RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat.Biotechnol. 31, 233–239.
- Hsu, P.D., Scott, D.A.,Weinstein, J.A., Ran, F.A., Konermann, S., Agarwala, V.,Li, Y., Fine, E.J., Wu, X., Shalem, O., et al. (2013). DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. Published online July 21, 2013.
- Ran, et al. (2013). Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 154, 1380.

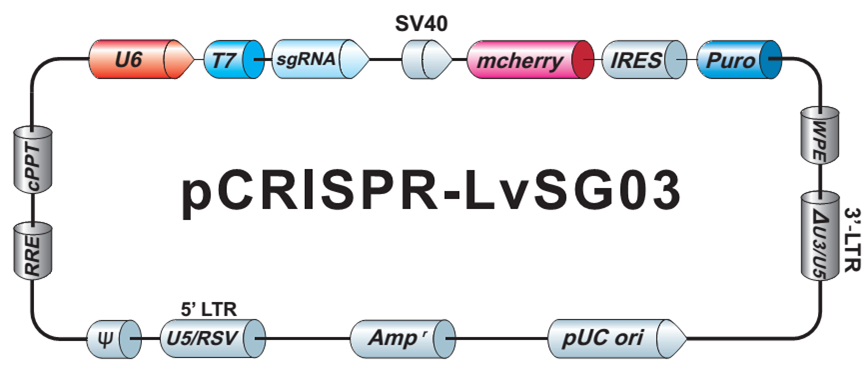
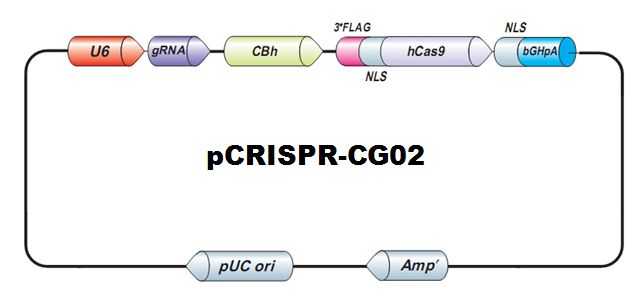
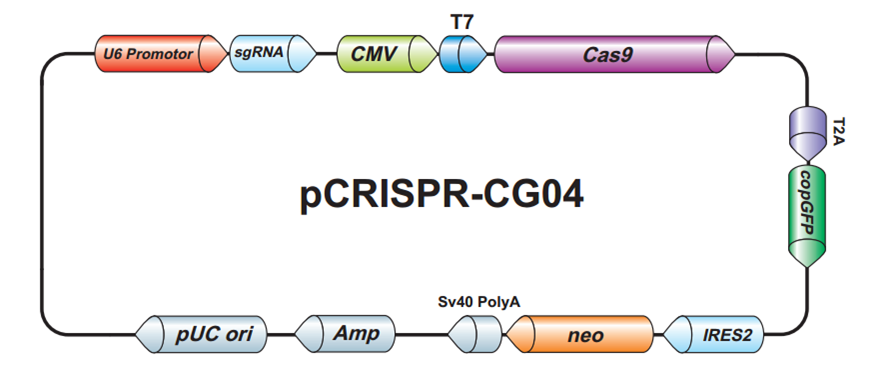
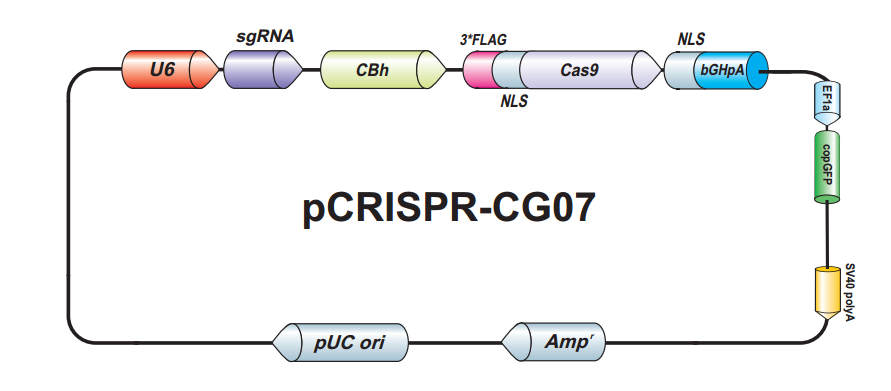
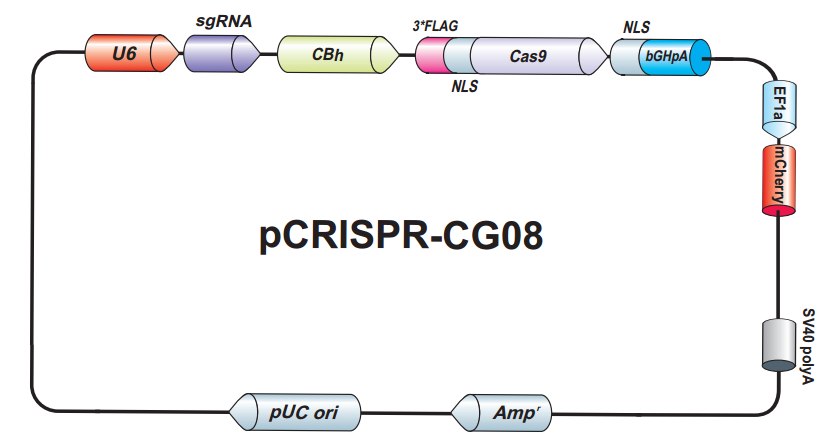
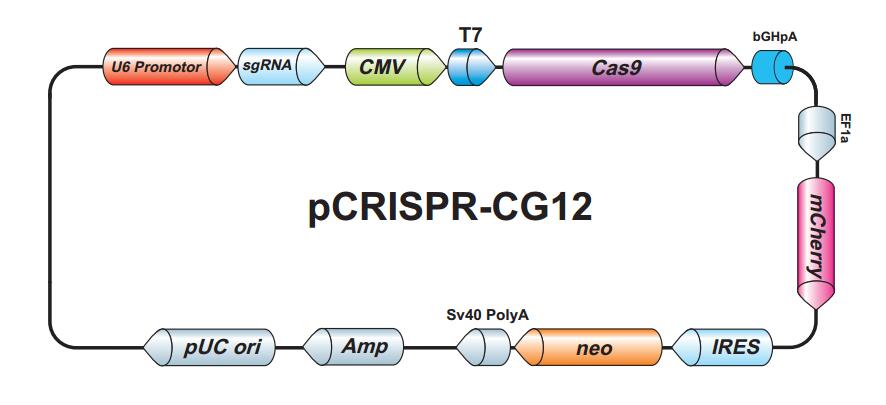

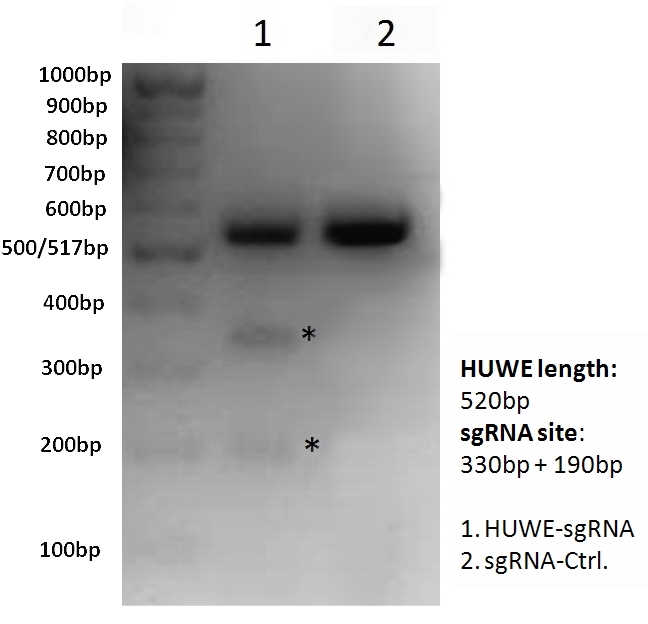
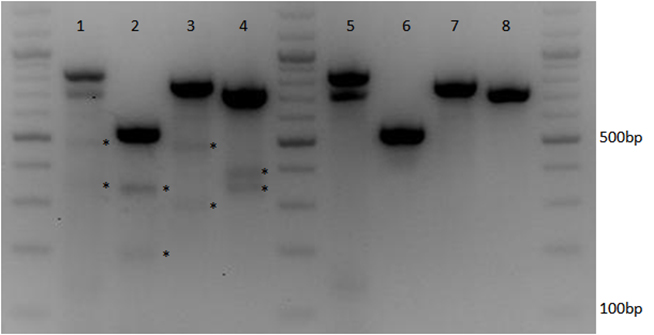





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}